


№9, 2007 р
© Алифанов В.Р.
Ящірки в епоху динозаврів
В.Р. Алифанов
Володимир Рудольфович Алифанов, кандидат біологічних наук, старший науковий співробітник Палеонтологічного інституту РАН.
Релікти юрського періоду
На тлі сучасного розмаїття ящірок, що досягає декількох тисяч видів, палеонтологічний літопис виглядає бідно. І не дивно! Останки невеликих тварин зазвичай швидко руйнуються і рідко зберігаються. Проте завдяки знахідці декількох дрібних щелепних кісток в нижньокрейдових відкладах Англії в середині XIX ст., Тобто на зорі розвитку палеонтології хребетних, з'ясувалося, що ящірки були сучасниками динозаврів. Але ще раніше з'явилася інформація про гігантських водних ящірок - мозазаври, що мешкали в морях на останньому етапі мезозойської ери.
З часу першої знахідки на території Голландії в 1780 р особливості способу життя і спорідненості мозазавров не раз обговорювалися в середовищі фахівців. У їх числі були такі відомі палеонтологи, як Ж.Кювье, Р. Оуен, Е. Коп, О.Марш і Л.Долло. В результаті вдалося встановити, що виникнення і розквіт морських ящірок - всього лише яскравий епізод складного і тривалого історичного розвитку в цілому наземної групи, яка, як зараз достеменно відомо, з'явилася на арені еволюції в середині юрського періоду, тобто близько 170 млн років тому.
Початок поширення ящірок обумовлено рядом причин, в тому числі тим, що протягом мезозойської ери на планеті був відносно рівний і теплий клімат, без оледенений і зі слабо вираженою широтной зональностью. Це дуже важлива обставина не тільки для ящірок, але і для всіх плазунів, оскільки їх активність, на відміну від птахів і ссавців, залежить від зовнішнього тепла. З одного боку, подібна фізіологічна особливість дозволяє економно витрачати енергію, що надходить з їжею, а з іншого, ставить в залежність від добових або сезонних температур. Чим холодніше або чим більше температурні коливання, тим важче гадів закріпитися на тій чи іншій території. Ось чому в мезозої вони легко розселялися у всіх напрямках, а сучасна Герпетофауна зосереджена головним чином в субтропічних і тропічних широтах.

Скелет хайнозавр - одного з багатьох видів мозазавров.
Ці морські ящірки були найстрашнішими хижаками морів в кінці мезозою.
Деякі з них, як хайнозавр, досягали 15-17 м в довжину.
Багато в чому еволюційний зліт ящірок пов'язаний з рядом фізіологічних і морфологічних новопридбаних. Найбільш важливі з них стосувалися будови черепа. В результаті верхня щелепа ящірок придбала рухливість, а щелепної апарат в цілому отримав можливість амортизувати ривки видобутку, перехоплювати її для максимально вигідного прикладання сили щелеп і швидко проштовхувати в глотку. Розрахунки показують, що рухливий, або кінетичний череп (у ящірок черепна конструкція називається амфікінетіческой через наявність двох поворотних зон в даху черепа) максимально ефективний, якщо видобуток невелика і не вимагає будь-якої спеціальної обробки щелепами. За цими показниками для ящірок найкращим чином підходять комахи [ 1 ], Що стали виключно численними на початку мезозою.
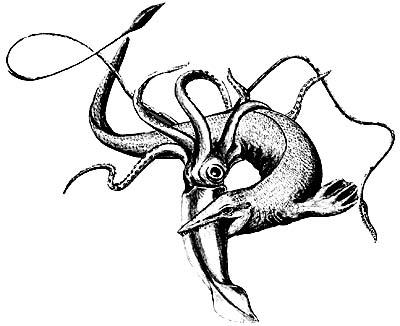
Сцена полювання мозазавра.
Їжею морським ящірок служила невелика риба,
але вони вступали в сутички і з різноманітними ворогами,
в тому числі і з гігантськими кальмарами.
примхи літописі
Науковий інтерес до ящірок проявився на рубежі XVIII-XIX ст. Тоді фахівці, знайомлячись з їх різноманітністю, спиралися головним чином на зовнішні характеристики. У другій половині 19-го століття, коли почали поширюватися еволюційні ідеї, виникла необхідність створити филогенетическую класифікацію ящірок, для чого потрібні були дані порівняльної анатомії. На цьому етапі палеонтологія не зробила помітного впливу, поставивши в число основних лише питання про походження і родинних зв'язках мозазавров.
В кінці XIX-початку XX ст. з'явилося відразу кілька розгорнутих класифікацій. Однак в основу сучасної картини лягла система американського вченого Ч.Кемпа [ 2 ]. У ній фігурують назви прийнятих і зараз великих систематичних утворень (Інфраотряд): сцінкоморфи (сцинки і їх родичі), ангвіморфи (варани, ядозуби, веретеніци і їх родичі), геккоти (гекони і чешуєног) і ігуану (агами і ігуани).
Кемп вказував на близьку спорідненість сцінкоморф і ангвіморф, а також геккот і ігуану, причому другу пару вважав архаїчніше першої. В останні десятиліття ці уявлення не раз піддавалися переосмисленню. В результаті геккоти були зближені з сцінкоморфамі і ангвіморфамі в складі загальної групи Сцинки-геккономорф, а статус найбільш близьких до предковому типу ящірок збережений тільки за ігуану [ 3 , 4 ]. Але останній висновок не безперечно. Йому, наприклад, суперечать дані палеонтологічного літопису, в якій першими про себе заявляють Сцинки-геккономорфи.
Втім, у палеонтологічному літописі є свої примхи. Наприклад, ще недавно побутувало уявлення про кайнозойської розквіті ящірок на тій підставі, що переважна кількість їх викопних знахідок виявлено у відкладеннях відповідного віку. Однак в останні десятиліття все виразніше став вимальовуватися мезозойский підйом різноманітності групи. Цьому сприяло вивчення численних матеріалів з континентальних відкладень пустелі Гобі, різноманітних систематично і унікальних по схоронності. У Монголії копалини ящірки збиралися і оброблялися учасниками кількох міжнародних палеонтологічних експедицій: Польсько-Монгольської (1964-1972 рр.), Радянсько-Монгольської (з 1969 р) і Монголо-Американської (з 1992 р).
Але перші збори були зроблені на початку 20-х років XX ст. експедицією Американського музею природної історії (Нью-Йорк) на верхнемелового місцезнаходження Баин-Дзак. У пресі їх представив американський палеонтолог Ч.Гілмор [ 5 ]. З кількох фрагментарних зразків найбільш примітним виглядає макроцефалозавр (Macrocephalosaurus), череп якого виявився на рідкість, навіть для сучасних форм, значного розміру - більше 10 см в довжину (цим і пояснюється його назва: большеголовая ящірка). Цікава ще одна деталь: на передніх кінцях верхніх щелеп макроцефалозавра були великі іклоподібні зуби, як у сучасних агам. Ця та деякі інші особливості схилили вченого до висновку про зв'язок макроцефалозавра саме з агамообразнимі ящірками.
Збори інших експедицій істотно розширили уявлення про різноманітність копалин ящірок Азії, але одночасно оголили нові проблеми. Зокрема, виявилося, що багато крейдяні форми не піддаються однозначному систематичного визначення. Так, польський палеонтолог А.Сулімскій припустив, що встановлений ним по повному черепу адамізавр (Adamisaurus) знаходиться в тісній спорідненості з агами [ 6 ]. Дійсно, такий висновок можна зробити, спираючись на будову, скажімо, кісток нижньої щелепи. Однак через кілька років Сулімскій вирішив, що схожість адамізавра і агам поверхнево, а гіпотезу спорідненості викопної форми необхідно переглянути на користь сцінкоморф [ 7 ]. Польський палеонтолог описав також кілька нових видів макроцефалозавров [ 8 ]. Як виявилося, їх "закликовие" зуби мали лопаткообразную вершину, розділену на численні додаткові зубчики, як у деяких сучасних ігуан. Але ці деталі дослідник вважав незначними і, як і адамізавра, відніс макроцефалозавров до сцинкоподібні ящірок.

Череп адамізавра (Adamisaurus magnidentatus).
За формою зубів він нагадує деяких Сцинки-геккономорфних ящірок,
але будовою нижніх щелеп близький до агамідам.
Зразок з фондів Палеонтологічного інституту (СІН).
Проблему спорідненості адамізавра і макроцефалозавров спробував вирішити американський палеонтолог Р.Естес [ 9 ]. Йому належить ідея включити цих копалин ящірок до складу тейид (Teiidae) - сімейства, представники якого нині населяють головним чином Південну Америку, а в крейдяне час були поширені по всій Америці. Цікаво, що положення цієї групи, що складається з двох підгруп (макро- і мікротейін), в системі ящірок точно не встановлено. Деякі систематики допускають, що макротейіни, яких ще називають американськими варанами, споріднені ігуану. Однак загальноприйнята, але не має очевидних морфологічних доказів точка зору вказує на приналежність цієї групи сцінкоморфам.
Протиріччя в судженнях фахівців мають свої причини. Головна з них в тому, що діюча класифікація, створена переважно на підставі сравнітельноанатоміческого вивчення сучасних форм, не завжди відповідає древньому різноманітності. Разом з тим саме по знахідкам копалин тварин вдається розкрити помилки, неточності і протиріччя багатьох сформованих раніше поглядів. І хоча виклад природної історії ящірок в наш час вже важко уявити без урахування палеонтологічного матеріалу, але сказати, що він лежить в основі сучасних уявлень про еволюцію цієї групи плазунів, ще не можна.
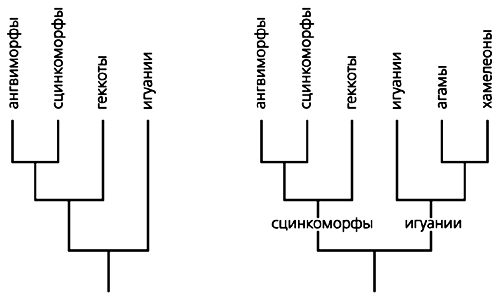
Варіанти родинних зв'язків основних підгруп ящірок. Зліва - варіант, запропонований Р.Естесом і ін. [ 4 ], Справа - автором [ 14 ].
Ігуани і історична зоогеография
Відкриття в складі комплексів позднемеловой тетрапод Азії ігуан (Iguanidae) сучасного типу для багатьох герпетологів стало несподіванкою [ 10 ]. Справа в тому, що в даний час ящірки цієї групи поширені в основному в Північній і Південній Америках. Правда, вони живуть ще на Мадагаскарі (три види) і на островах архіпелагів Фіджі і Тонга (один вид), що здається винятком із загального правила. Саме ж правило говорить: ігуани живуть в Новому Світі, а їх найближчі родичі агами (Agamidae) - в Старому Світі (крім Мадагаскару) і Австралії. Іншими словами: там, де мешкають ігуани, немає агам.
Встановленням і поясненням закономірностей розселення тварин у минулому займається історична зоогеография. Можливості цієї дисципліни ще далеко не вичерпані, про що можна судити по різночитання гіпотез, що пояснюють походження сучасної фауни Австралії з її великою кількістю ендемічних (тобто мешкають тільки на даній території) видів сумчастих ссавців. Якщо прийняти, що більшість видів плацентарних ссавців з'явилося на цьому оточеному з усіх боків морями південному материку разом з людиною або трохи раніше, то можна зробити деякі цікаві висновки. Один з них: центр походження сумчастих територіально був відокремлений від центрів походження інших груп ссавців. І наступний висновок: австралійська фауна відокремилася перш, ніж почалося розселення плацентарних.
За уявленнями ряду геологів і біологів, Австралія перетворилася на острів в крейдяне час, понад 120 млн років тому, після чого почала рухатися на північ. До цього в тріасовому періоді, близько 220 млн років тому, материк ще був частиною надконтиненту Пангеї, а потім входив до складу Гондвани, достовірно включала Австралію, Антарктиду і Південну Америку. На півночі розташовувався інший суперконтинент - Лавразия, куди входили Північна Америка, Європа і Палеоазія. Мабуть, Гондвана і стала батьківщиною сумчастих. Плацентарних ж дісталася Лавразия. Вона, до речі, проіснувала недовго - на кордоні юрського і крейдяного періодів розділилася на кілька обмежених морями блоків.
Протягом мезозою рівень моря постійно підвищувався (епоха трансгресії), що в другій половині крейдяного періоду призвело до ізоляції всіх великих ділянок суші. Єдино достовірне велике зоогеографічне подія того часу - обмін фаун Палеоазіі і Північної Америки через Берінгійской міст. Зазвичай вважається, що міст діяв протягом більшої частини пізньої крейди, проте є уявлення і про його короткостроковому існування. Остання точка зору заснована на тому, що позднемеловой фауни Азії і Північної Америки включають ендемічні групи, які нерідко формуються в умовах територіальної ізоляції. З урахуванням цієї обставини не можна виключити, що вперше азіоамеріканская зв'язок почала діяти на рубежі раннього і пізнього крейди.
Однак формування Берінгіі не пояснює всіх особливостей поширення наземних тварин в крейдяне час. Наприклад, одні групи динозаврів (тираннозаврид, гребнеголовие гадрозаври, вищі рогаті динозаври) заселяли Палеоазію і Північну Америку, а інші (плоскоголові гадрозаври, альваресзаври, анкілозаври з сімейства нодозавр) - ще й Південну Америку. Відсутність деяких типових для північних континентів груп динозаврів в Південній Америці можна пояснити двома способами: або неповнотою палеонтологічного літопису, або припущенням ще одного крупного зоогеографічний події.
Їм могла бути міжамериканська фауністична зв'язок, яка якщо мала місце, то до початку формування Берінгіі. Фактично можна припустити почерговість зв'язків Північної Америки: спочатку з Південною Америкою, а потім з Палеоазіей. Саме такий хід подій дозволяє зрозуміти, чому ареал азіатських за походженням динозаврів не розширені далі Північної Америки, тоді як їх неазіатського групи освоїли не тільки Південну або Північну Америку, а й Палеоазію.
Але повернемося до ігуанам. Розгадати таємницю їх розселення зовсім не просто. Р.Естес вважав, що у цієї групи виключно гондванські коріння. Пізніше вона зникла в Африці та Австралії, але збереглася в Південній Америці, де також виявлені її позднемеловой форми. На думку Естес, саме з території цього континенту на кордоні мезозою і кайнозою ігуани проникли в Північну Америку - імовірно на рослинних плотах.
Однак відкриття ігуан в позднемеловой відкладеннях Азії підриває логіку Естес. Тепер можна допустити поширення групи ні з півдня на північ, а в протилежному напрямку, не в кінці пізньої крейди, а на початку, і не на плотах, а по суші. Не зайвим буде зауважити, що розселятися за допомогою плотів могли б окремі види дрібних наземних тварин, в тому числі ящірки, але ніяк не динозаври. Для цих досить великих тварин такий спосіб явно не підходить. Так чи інакше, але батьківщиною ігуан могли стати і Північна Америка, і близька до неї Палеоевропа. Не можна скидати з рахунків і Африку, за деякими уявленнями, ще в юрський час втратила зв'язок з південною групою континентів і встановила фауністичне повідомлення з західною частиною Лавразии вже після ізоляції Палеоазіі [ 11 , 12 ]. В цьому випадку батьківщиною ігуан могла стати і Афроамерія, що об'єднувала через Європу Африку (з Мадагаскаром) і Північну Америку, а їх проникнення (в супроводі деяких груп американських динозаврів) на територію Палеоазіі сталося вдруге через Берінгійской міст. Це так ще й тому, що в Азії раннемелових ігуани не виявлені.
Палеонтологічні дані відкривають можливість по-новому поглянути і на заселення ігуанами о-вов Фіджі і Тонга, куди, як вважалося, вони потрапили на рослинних плотах з Південної Америки. Тепер припустимо припустити, що ігуани проникли на архіпелаги з Південно-Східної Азії, яка, судячи зі знахідок динозаврів, в крейдяне час була півострівний частиною палеоазіатской суші.
Отже, згадане правило виглядає результатом досить випадкового поєднання ряду історично сформованих обставин. Втім, без конкуренції між агам і ігуанами все ж не обійшлося.
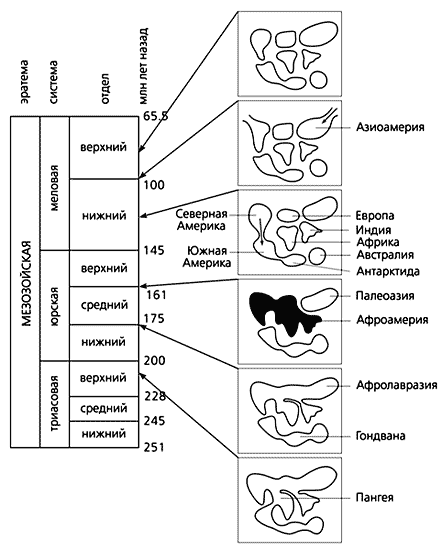
Реконструкція послідовності континентальних зв'язків в мезозої по фауністичним даними [ 11 , 12 ].
Афроамерія (виділена чорним), можливо, була ареалом первинного поширення ігуан
(кольоровими стрілками вказані головні напрямки їх розселення в крейдяне час).
вершина еволюції
На початку пізньої крейди центральна частина Палеоазіі представляла собою велику низовина, куди вітер і вода зносили великі маси піску. У сезони дощів тут з'являлося безліч тимчасових водойм, береги яких швидко заростали. На мілководдях і на березі численними ставали безхребетні: черви, рачки, комахи і їх личинки. Ними годувалися дрібні ссавці, птахи та ящірки. Однак ці хребетні і самі легко могли потрапити в щелепи хижих динозаврів - тероподів, серед яких переважали дромеозаврів, що мали, судячи з новітніми даними, птіцеообразний вигляд. З них більше за інших відомий двометровий велоцираптор (Velociraptor). У нього, як і у всіх його найближчих родичів, була невелика голова, довгі і вузькі щелепи, великі передні лапи і збільшений в розмірі кіготь на другому пальці стопи.
Тому, що тероподи не втрачали нагоди схопити і проковтнути ящірку, є незаперечний доказ - плита літографських сланців, на поверхні якої видно практично повний скелет компсогната (Compsognathus) із залишками баварізавра (Bavarisaurus) в області шлунка. Трапеза динозавра, що жив в центрі Європи, сталася в кінці юрського періоду (близько 150 млн років тому).
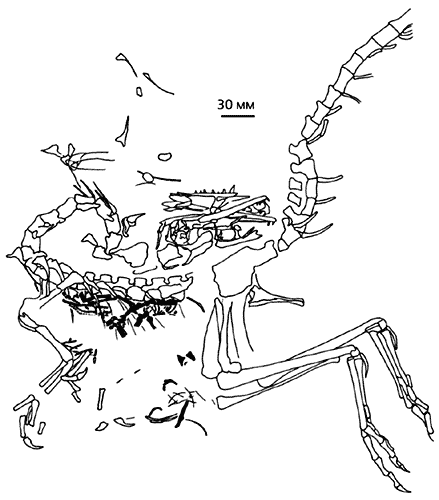
Скелет динозавра компсогната з верхнеюрских сланців Баварії
із залишками ящірки (виділені чорним кольором) баварізавра в області шлунка [ 16 ].
Існувалі й Такі ящіркі, зустрічей з Якими велоцираптори Унікал. Це Великі двометрові Естес (Estesia). З ними легко могли впоратися хіба що тираннозавра. Але динозаври-гіганти цуралися Гобійського низин, затоплюваних в сезон дощів, залишаючи на вершині харчової піраміди великих вараноідних ящірок. Вони, звичайно, програвали в швидкості більш маневреним і активним тероподам, але міць і здатність плавати дозволяла їм успішно конкурувати за простір і харчові ресурси.

Силуети варана Естес (зліва) і динозавра велоцираптора.
Міць і здатність плавати дозволяли великим варанам
успішно конкурувати з динозаврами за простір і харчові ресурси.
Зараз серед ящірок на положенні домінуючих хижаків знаходяться тільки варани о.Комодо, довжина тіла яких доходить до 3 м. Приблизно мільйон років тому в Австралії ту ж роль грали семиметрові мегаланія. Гігантизм нетиповий для ящірок, але може сформуватися при якихось особливих обставин. Судячи з умов проживання комодского дракона і мегаланія, можна припустити, що збільшення розмірів стимулюється кількістю їжі і відсутністю конкуренції.
Говорячи про варанів, не можна не згадати про те, що ще з часів Кемпа вони вважаються предками змій. Ця гіпотеза, що отримала назву платінотной (від Platynota - назви вараноідних ящірок в класифікації Кемпа), заснована на окремих ознаках подібності двох груп. У числі таких рис згадуються, наприклад, довгий і рухливий мову; великі капсули якобсонова органу, відповідального за хеморецепции; гострі зуби; додаткове рухливе зчленування гілок нижньої щелепи; великі виростків (виступи) тіл хребців. Насправді всі ці особливості разом властиві лише представникам сучасних варанід (Varanidae), тоді як у їхніх викопних і сучасних родичів схожість зі зміями практично не виражено. Швидше, всі перераховані ознаки відбулися паралельно в зв'язку з переходом варанів і змій на харчування невеликими хребетними. До речі, щелепний апарат вараноідних зберігає типові для ящірок особливості, хоча забезпечений порівняно більш потужною мускулатурою. Зовсім інакше йде справа з щелепним апаратом змій.

Грудні хребці сучасного сірого варана (зліва), що досягає 1.5 м в довжину,
і викопної вараноідной ящірки (Cherminotidae), перевищувала 2 м (верхня крейда Монголії).
Прим. СІН, №4216 / 206.
Харчова спеціалізація більшості змій спрямована на проковтування великої здобичі. Це забезпечується високою взаємною рухливістю і розтяжністю деяких внутрішньочерепних з'єднань, одночасно ведуть до втрати сили щелеп. Ось чому багато змій, щоб придушити опір жертви, попередньо її душать або кусають, впорскуючи отруту. Таким чином, спеціалізація щелепного апарату змій і ящірок різна, і лише по ній зв'язати дві групи ставленням предки-нащадки важко.
До речі, змії, що з'явилися в раннемеловое час в Північній Африці, стали звичайним елементом комплексів хребетних тільки в кайнозої. У мезозойських відкладеннях Азії поки не знайдено жодної викопної змії. Дана деталь чітко ілюструє, що крейдяні ящірки не відчували протидії з боку їх найнебезпечніших конкурентів.
пора розквіту
Найдавніша азіатська ящірка - чангетізавр (Changetisaurus) з среднеюрских відкладень Киргизії. Її відносять до парамацеллодідам (Paramacellodidae), вимерлого сімейства сцінкоморф. Парамацеллодід знаходять в позднеюрских, іноді в раннемелових відкладеннях всіх північних континентів, як і дорсетізаврід, ймовірно, належали ангвіморфам.
Численними ящірки стають у другій половині раннього крейди разом з початком періоду потепління, яке відбулося приблизно 120 млн років тому. Зміни відбилися і на розмаїтті інших груп хребетних, в тому числі динозаврів. Про розквіт ящірок того часу найкраще можна судити по знахідках з монгольського місцезнаходження Хобур, розташованого поблизу південно-східного краю Хангайский хребта. У комплексі представлений десяток родин, в основному вимерлі групи Сцинки-геккономорф. Є в ньому парамацеллодіди і дорсетізавріди. Єдина достовірно визначена сучасна група в Хобуре - гекони [ 13 ].

Залишки раннемелових ящірок з місцезнаходження Хобур.
В основному це розрізнені елементи скелета,
серед яких переважають щелепні кістки Сцинки-геккономорф.
До початку пізньої крейди деякі колишні сімейства ящірок вимерли, але з'явилося багато нових груп, частина з яких мігрувала з Північної Америки через Берінгійской міст. Тільки на півдні Монголії з декількох верхньокрейдяних місцезнаходжень відомо 18 сімейств. Такого не знає жоден регіон ні в давнину, ні в сучасності. Мабуть, цей показник, як ніякий інший, свідчить про справжній часу розквіту ящірок.
Основне різноманітність позднемеловой ящірок древньої Азії становили ігуану. Тут були представлені не тільки всі филогенетические гілки цієї групи, але і близький до сучасного набір життєвих форм. Так, макроцефалозаври, що належали, судячи з будови нижньої щелепи, агам-хамелеоновому стовбура ігуану, нерідко нагадували сучасних великих ігуан, але володіли "іклами". У деяких з великоголових ящірок, як, наприклад, у апрізавра (Aprisaurus), їх було кілька пар. Однак інші макроцефалозаври зовні схожі на макротейін. Такий був джадохтозавр (Dzhadochtosaurus). Власне макротейіни теж виявлені в комплексах древньої Азії, але ці ящірки були там рідкісні.
Слід сказати і про велику різноманітність так званих пріскагам, які зовнішнім виглядом і будовою зубної системи нагадували агам з характерними для них "іклами" і сплощеними субтреугольнимі закликовимі зубами. Однак найближчі сучасні родичі пріскагам - хоплоцерк (Hoplocercus) і моруназавр (Morunasaurus) - живуть у Південній Америці і вважаються ігуанами. А ось у справжнього родича агам, ізодонтозавра (Isodontosaurus), зуби позбавлені явних ознак подібності з зубами сучасних представників, в тому числі і за характером прикріплення.
Зазвичай зубну систему агам і хамелеонів відносять до акродонтному типу, при якому зуби займають на щелепних костях верхушечное положення. Іноді ящірок цих груп навіть називають акродонтнимі ігуану. Особливий статус акродонтності надав американський вчений Е. Коп ще в XIX в. З тих пір термін міцно устоявся в літературі як один з трьох основних способів прикріплення зубів у наземних хребетних поряд з текодонтние (зуби сидять в поглибленнях - теках, характерний, наприклад, для крокодилів) і плевродонтним (зуби приростають до щелепи збоку, як у більшості ящірок ). Однак критичний погляд на особливості будови зубної системи акродонтних ігуану показує, що зуби, які закладаються на ембріональних стадіях онтогенезу, міцно зростаються з щелепними кістками і перестають рости. У міру збільшення індивідуальних розмірів тварин ембріональні зуби стають мініатюрними, зберігаючи своє положення поблизу крайок щелеп, тобто стають акродонтнимі. Однак початково їх тип прикріплення плевродонтний, як у "іклів" (якщо вони виражені) або постембріональному виникли зубів. Таким чином, типологічна особливість зубної системи акродонтних ігуану виглядає перебільшеною, що, як з'ясовується, призвело і до недооцінки різноманітності цієї групи, в тому числі викопного. До речі, у ізодонтозавра немає "іклів", ембріональні і постембріональний зуби майже не відрізняються один від одного морфологічно, а ось процес змінності явно обмежений.
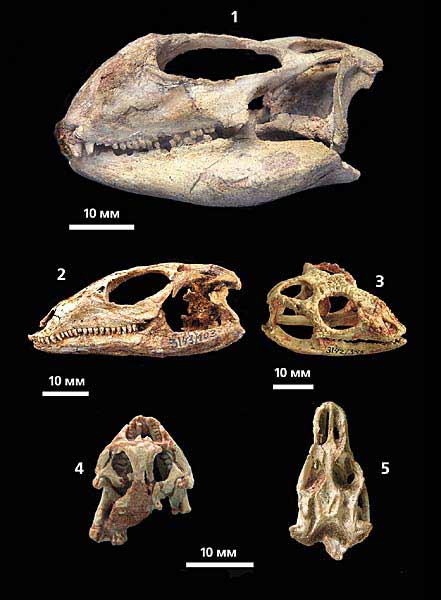
Черепа деяких позднемеловой ігуаніевих ящірок Монголії:
1 - апрізавра (Aprisaurus bidentatus, прим. ПІН №3142 / 302);
2 - джадохтозавра (Dzadochtosaurus giganteus, прим. ПІН №3143 / 103);
3 - хамелеогната (Chamaeleognathus iordanskyi, прим. ПІН № 3142/345);
4 - ізодонтозавра (Isodontosaurus gracilis, прим. Ін-ту палеобіології Польської АН, MgR II / 39);
5 - параварана (Paravaranus angustifrons, прим. ПІН № 4487/15).
Нові палеонтологічні знахідки в крейдяних відкладеннях Монголії показують, що проблема різноманітності древніх і копалин ігуану стає виключно актуальною. І тут можливі деякі несподівані рішення і підходи. Наприклад, розгляду вимагає грунтовно забута і випала з обговорення гіпотеза про спорідненість з ігуану мозазавров. Поки переважає уявлення про зв'язок мозазаврообразних ящірок з вараноіднимі ящірками і зі зміями, яка апелює до деяких загальних і виникли вдруге ознаками черепа.
До речі, до мозазаврообразним ящірок з крейдяних відкладень Монголії відноситься параваран (Paravaranus). Як і у типових морських ящірок, у нього відзначаються подовжені носові отвори і вузька лобова кістка. Зближують параварана з мозазаврідамі і деякі загальні риси в будові кісток даху черепа і піднебіння. Однак на відміну від морських ящірок параваран відрізнявся мініатюрними розмірами і, швидше за все, вів наземний спосіб життя.
Отже, різноманітність мезозойських ящірок виглядає досить переконливо. Найважливішим підсумком його вивчення стало уявлення про виключне видовому багатстві ігуану в минулому і про те, що таксономічний ранг самої групи не поступалася такого Сцинки-геккономорф [ 14 ].

Реконструкція черепа (вид зверху і збоку) сінеоамфісбени (Sineoamphisbaena hexatabularis).
На відміну від інших амфісбен у неї є Заочноямкові (зд) і верхні скроневі дуги (ВВД).
Чорним кольором показані носові отвори (але), орбіти (о) і верхні скроневі вікна (вво).
Говорячи про крейдяних ящірках древньої Азії, не можна обійти увагою сінеоамфісбену (Sineoamphisbaena), описану в літературі по знахідкам в верхньокрейдяних відкладеннях Внутрішньої Монголії (Китай) [ 15 ]. Широка кісткова межглазничного перегородка, велика мозкова капсула, роздвоєний потиличний мищелок і укорочена нижня щелепа і ряд інших ознак зближують цю викопну форму з амфісбенамі, або амфісбени. Сучасні амфисбени ведуть потайний спосіб життя - мешкають в норах, які риють головою. Мабуть, з цим пов'язані такі перетворення, як втрата кінцівок, редукція деяких кісток даху черепа. Однак на відміну від раннекайнозойскіх і сучасних амфісбен, у сінеоамфісбени були добре розвинені передні кінцівки (задні не виявлені), а елементи верхніх скроневих дуг нагадували ці ж деталі будови у ящірок, хоча і мали деякі обмежують амфікінетізм особливості. І те, і інше свідчить про архаїчності викопної форми, яка ще не досягла досконалості в роющей спеціалізації. Схоже, що в образі сінеоамфісбени проявляються риси безсумнівного спорідненості амфісбен і ящірок. Але включати перших до складу друге немає необхідності.
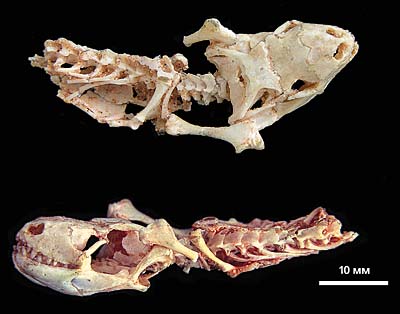
Череп і передня частина скелета славойі (Slavoia darevskyi), архаїчної сцінкоморфной ящірки з верхньокрейдяних відкладень Монголії.
У славойі парадоксально поєднуються мініатюрні очниці і добре розвинені кінцівки. Прим. ПІН № 4487/14.
Слід зауважити, що в різних філогенетичних стовбурах Сцинки-геккономорф існують форми, які відтворюють ті чи інші морфологічні особливості амфісбен. Нерідко вважається, що подібного роду подібності виникли вдруге, а первинної була наземна спеціалізація. Однак не виключено, що початково ящірки вели потайний спосіб життя. Надалі одні їх групи освоїли підземні біотопи, втративши кінцівки і придбавши здатність рити нори головою, а інші пристосувалися до наземного життя, удосконаливши способи пересування і розвинувши зір і слух. Прикладом "стародавнього" типу може служити сцінкоморфная славойя (Slavoia), яка в пізньому крейдяному періоді Монголії була найбільш масовою. У цій мініатюрній ящірки поєднується, як здається, несумісне - незначні розміри очних ямок (тобто крихітні очі), дозволені підземним жителям, і добре розвинені кінцівки, такі необхідні для наземного пересування.
В епоху змін
Принаймні за кілька мільйонів років до кінця мезозою різноманітність азіатських ящірок скоротилося більше ніж в чотири рази. Тоді число сімейств знизилося з 18 до чотирьох. Був ще один і більше пізній криза, яка виявляється за складом раннекайнозойскіх груп: крейдяні коріння тільки у однієї з них. Таким чином, неухильне зростання різноманітності протягом юри і крейди несподівано змінився швидким падінням.
Ключем до розуміння головної причини кризи може послужити раннекайнозойская історія групи.
На початку кайнозою в центральній частині Азії ящірки знову виявилися «на підйомі», але за рахунок відсутніх за старих часів сімейств. Мабуть, їх заселення відбувалося вдруге, з сусідніх територій. Судячи з аналізу систематичного складу раннекайнозойскіх комплексів, процес протікав неодноразово, а хвилеподібно протягом палеогенового періоду. Перша хвиля прийшла з північного сходу, через Берингову, з Північної Америки і пов'язаної з нею на початку кайнозою Європи. Друга - з півдня, з Індії, яка приєдналася до Азії після крейдяний ізоляції. Третя - західна, або європейська, - виникла після того, як Європа втратила зв'язок з Північною Америкою, а відділяла Європу від Палеоазіі в другій половині мезозою і на початку кайнозою Тургайське море висохло. Завдяки північно-східній хвилі з'явилися агами, близькі до сучасної Агама-метелику (Leiolepis). З південного хвилею прибутку неотруйні змії, варани сучасного типу і агами, схожі з сучасним шипохвостов (Uromastyx). Західна хвиля принесла в Центральну Азію ящірок тих пологів і сімейств (в тому числі справжніх ящірок - Lacertidae), які зараз звичайні в помірних широтах Євразії.
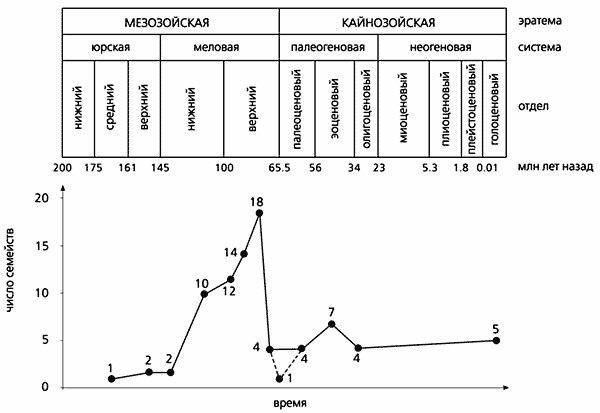
Динаміка розмаїтості сімейств копалин ящірок в мезозої і в кайнозої.
У кайнозої найбільше і найменше число сімейств збігаються з періодами найвищих і найнижчих палеотемператур.
Ймовірно, цим же пояснюються і мезозойські максимуми і мінімуми різноманітності.
Примітно, що залежні від умов навколишнього середовища ящірки в кайнозої, як і в крейді, поширювалися через високоширотних Берінгійской міст. Це пов'язано з черговим потеплінням, який привів в середині першої половини кайнозойської ери (в еоцені) до встановлення найвищого для того часу температурного максимуму. На тлі потепління число сімейств ящірок збільшилася з чотирьох до семи. Однак до кінця еоцену різноманітність центральноазіатських ящірок знову впало. Причина - добре відоме похолодання. Воно ж прямо або опосередковано призвело навіть до збіднення комплексів домінуючих в кайнозої ссавців. Швидше за все, позднемеловой криза азіатських ящірок і рептилій в цілому - теж наслідок одного або серії похолодань.
Як би там не було, але, на відміну від динозаврів, ящірки дожили до наших днів. Рятівними для них стали невеликі розміри, здатні забезпечити швидку зміну поколінь, полегшити пристосовність до різних біотопів і компенсувати втрати чисельності і різноманітності в післякризові часи. Ймовірно, саме цих якостей не вистачило динозаврам в епоху глобальних змін на рубежі мезозойської і кайнозойських ер.
Робота виконана в рамках програми Президії РАН 25 "Походження і еволюція біосфери" на 2005 р (підпрограма 2)
і підтримана грантом Президента РФ для Провідною наукової школи НШ-6228.2006.4.
література
1. Йорданський М.М. Еволюція комплексних адаптацій. Щелепної апарат амфібій і рептилій. М., 1990. ..
2. Сamp CL // Bull. Am. Mus. Nat. Hist. 1923. V.48. P.289-481.
3. Суханов В.Б. // Зоол. журн. 1961. Т.40. №1. С.73-83.
4. Estes R., de Queiroz K., Gautier J. Phylogenetic relationships within Squamata // Phylogenetic relationships within lizards families / Eds R.Estes, GKPregill. Stanford, 1988. P.119-281.
5. Gilmore CW // Bull. Amer. Museum Natur. Hist. 1943. V.81. P.361-384.
6. Sulimski A. // Palaeontol. Polonica. 1972. V.27. P.33-40.
7. Sulimski A. // Palaeontol. Polonica. 1978. V.38. P.43-56.
8. Sulimski A. // Palaeontol. Polonica. 1975. V.33. P.25-102.
9. Estes R. Sauria terrestria, Amphisbaenia // Handbuch der Palaeoherpetologie. Stuttgart, 1983. T.10A.
10. Borsuk-Bialynicka M., Alifanov VR // Acta Palaeontol. Polonica. 1991. V.36. №3. P.325-342.
11. Каландадзе І.І., Раутиан А.С. Система ссавців і історична зоогеография // Філогенетика ссавців. М., 1992. С.44-152.
12. Kalandadze NN, Rautian AS Historical zoogeography of terrestrial tetrapods and new method of global palaeogeographical reconstructions // Evolution of the Biosphere / Eds Rozanov A.Yu., Vickers-Rich P., Tassel C. Launceston, 1997. P.95- 98.
13. Алифанов В.Р. // палеонтолого. журн. 1999. №1. С.124-126.
14. Алифанов В.Р. Макроцефалозаври і ранні етапи еволюції ящірок Центральної Азії. М., 2000. ..
15. Wu X. -Ch., Brinkman DB, Russell AP // Can. J. Earth Sci.1996. V.33. №4. P.541-578.
16. Evans SE // N. Jb. Geol. Palдont. Abh. 1994. Bd.192. №1. P.37-52.


